{kind=link}
{kind=link}
{kind=link}

| Potentiels récepteurs, Potentiels post synaptiques excitateurs, potentiels daction : |
| Déclenchement, propriétés et rôle dans le traitement des informations sensorielles. |
Les cellules sont des éléments fermés et délimités par une membrane semi-perméable. La composition des milieux intra et extracellulaires sont différentes. Lorigine de la différence de potentiel provient de linégale répartition des ions entre les deux faces de la membrane. Le maintien de part et dautre de la membrane de deux milieux différents permet aux cellules de stocker de lénergie sous forme de différence de potentiel. Cest justement cette énergie que va utiliser la cellule lors dune stimulation.
I. Potentiels de récepteursCest au niveau des récepteurs sensoriels que se fait la première étape du codage de linformation. Ces récepteurs peuvent être relativement facilement exposés, et stimulés par un léger étirement du muscle quils innervent. Il est alors possible denregistrer lactivité de ces récepteurs, soit dans le corps cellulaire, soit le long de laxone, lors de lapplication dun étirement. Une telle stimulation provoque lapparition dune variation de la différence de potentiel transmembranaire, correspondant à une faible dépolarisation de la membrane. (Fig.1)
Cette variation de potentiel est qualifiée de potentiel de récepteur, et caractérise la première étape de codage des stimuli.
Comme toutes variations de la différence de potentiel transmembranaire observée au niveau dune cellule, celles-ci sont dues à louverture de canaux ioniques particuliers.
Si lon augmente lintensité de la stimulation, lamplitude du potentiel de récepteur augmente de façon proportionnelle (Fig.1-A). Il y a donc codage, à ce niveau, de lintensité de la stimulation en une amplitude de dépolarisation transmembranaire du récepteur. On parle alors de codage en amplitude.
Si on réalise deux stimulations successives de faibles amplitudes, on peut constater quil y a sommation des effets. Le second potentiel de récepteur vient en effet sadditionner à la dépolarisation déjà existante (Fig.1-B).
Si maintenant, on place lélectrode denregistrement le long de laxone, on peut alors constater quil devient impossible denregistrer ces potentiels dus à une stimulation. Ces phénomènes sont donc des phénomènes électriques locaux conduits de façon purement électrique.
Le potentiel de récepteur est donc un phénomène graduable, sommable, et non propagé.
En suivant le même protocole expérimental, si on augmente encore lintensité de la stimulation, on peut alors voir apparaître des potentiels daction sur la fibre sensorielle (Fig.1-C). A partir de ce moment, et si on continue à augmenter lintensité de la stimulation, le nombre de potentiel daction par unité de temps, c'est-à-dire leur fréquence, augmente également (Fig.1-D). Linformation, précédemment codée sous la forme dune variation damplitude de la différence de potentiel transmembranaire, est désormais codée sous forme dune variation de fréquence de potentiels daction.
Les potentiels daction ainsi formés sont propagés de façon régénératrice le long de la fibre sensorielle.
La majorité des récepteurs sensoriels présentent un phénomène dadaptation, il existe en réalité deux types fonctionnels différents de ces récepteurs, localisés sur des fibres musculaires différentes. Lors dune stimulation prolongée, il est en effet possible de distinguer deux types dactivités différentes. (Fig.2) :
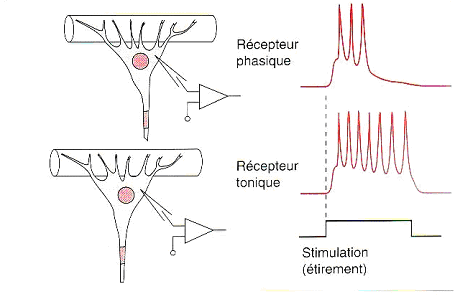
Les systèmes sensoriels phasiques ne peuvent renseigner le système nerveux central que sur la variation de stimulations et non sur leurs durées. A lopposé, les systèmes toniques permettent de coder la durée dune stimulation.
Les voies afférentes véhiculent linformation sensorielle vers le système nerveux central. Les messages nés au niveau des récepteurs sensoriels sont véhiculés par des fibres afférentes vers le système nerveux. Au niveau médullaire, par exemple, ces fibres pénètrent dans la moelle épinière par la racine dorsale, puis font relais dans différentes structures du système nerveux central. Il existe, au moins pour les principales modalités sensorielles, un relais au niveau du thalamus avant que les informations ne soient véhiculées vers des aires corticales spécifiques.
II. Potentiels dactionLintensité dune stimulation est codée sur les fibres sensorielles sous la forme dune fréquence de potentiels daction. Cette fréquence varie en fonction de la stimulation. Cette évolution de la fréquence en fonction de lintensité de la stimulation, est bien évidemment liée à la vitesse dadaptation du récepteur mis en jeu. (Fig.3).

Les potentiels daction napparaissent sur la fibre sensorielle que pour une stimulation minimale qualifiée dintensité seuil. Cette évolution de lactivité le long des fibres sensorielles permet de rendre compte de lévolution des sensations en fonction de lintensité dune stimulation.
Les potentiels daction ont deux origines possibles, il sagit de potentiels daction liés à des mouvements de sodium (Na+) et potassium (K+) ou de potentiels daction liés à des mouvements de calcium (Ca++). Nous nous intéresserons essentiellement aux potentiels daction sodiques.
La figure 4 représente une stimulation électrique.
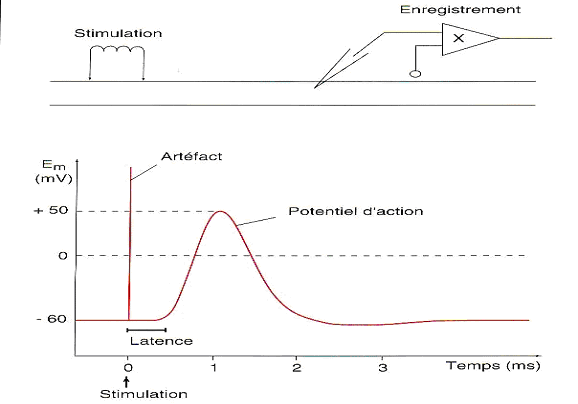
Cette dernière constitue un artefact suivi dun second phénomène (non électrique) puisquil apparaît après un temps de latence. Ce phénomène est la manifestation électrique de lactivité nerveuse. Cest ce phénomène qui est appelé Potentiel dAction.
Il a une amplitude de 120 mV et une durée de 2 ms. Il correspond à une variation de la différence de potentiel transmembranaire. Ceci suppose donc lexistence de courants électriques de part et dautre de la membrane de laxone.
Au sommet du potentiel daction la différence de potentiel est de +50 mV, nous observons quil est voisin du potentiel déquilibre des ions Na+ (équation de Nernst) :
On peut donc conclure que le potentiel daction correspond à une perméabilité momentanée de la membrane aux ions sodium. Lorsque la membrane atteint un certain niveau de dépolarisation, la probabilité douverture des canaux Na+ augmente, sa conductance aussi. Il apparaît alors un courant entrant de Na+ qui dépolarise la membrane. Lorsque le potentiel déquilibre est atteint il ny a plus de mouvement de Na+, sur la figure 5 nous sommes au sommet du potentiel daction.
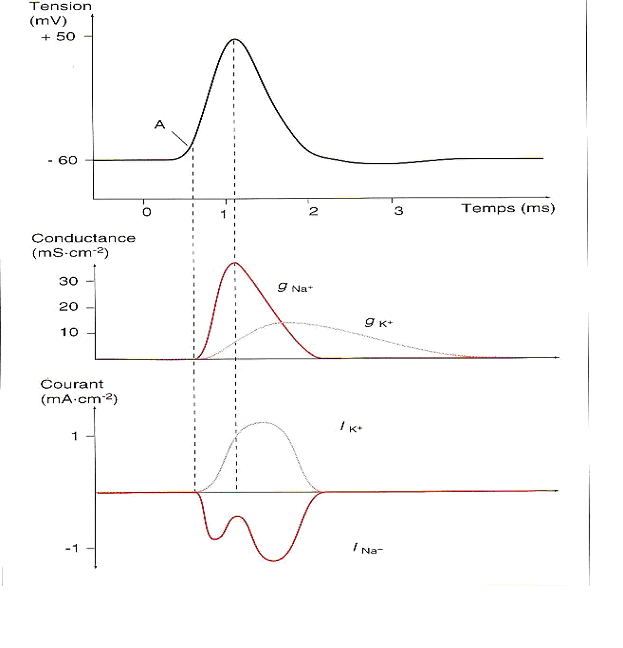
Pendant ce temps, la conductance au K+ augmente et la différence de potentiel transmembranaire séloigne du potentiel déquilibre de cet ion, donc sa force de diffusion augmente. Un courant K+ sortant apparaît donc. Ce dernier à un effet inverse par rapport au courant Na+ et permet de ramener rapidement la membrane à sa différence de potentiel de repos : cest la repolarisation. (Fig.5).
Ce sont des protéines appelées canaux ioniques spécifiques qui permettent le passage des ions du compartiment à lautre. Ces canaux sont fermés au repos et sont susceptibles de souvrir lorsque la membrane est dépolarisée. Ces canaux souvrant lors dune variation de la différence de potentiel transmembranaire, sont qualifiés de canaux sensibles à la tension ou tension dépendants. Les courants liés à louverture des canaux sensibles à la tension correspondent à des mouvements dions ayant deux cinétiques différentes : une rapide et une lente. Le courant rapide est un courant entrant de Na+ tandis que le courant lent correspond à une sortie de K+.
(On a pu le vérifier grâce notamment à des drogues telles que la Tetrodotoxine qui bloque les canaux Na+, et le Tetra-Ethyl-Ammonium qui bloque les canaux K+).
Les canaux jouent le rôle de porte. Ils peuvent être ouverts ou fermés. (Fig.6).

Ils souvrent sous leffet de la stimulation puis se referment et entrent dans une période dinactivation. Cela permettra au potentiel daction de ne pas revenir en arrière et de progresser dans le même sens.
Le potentiel daction est régénéré de proche en proche le long de la membrane. La vitesse de propagation du potentiel daction est beaucoup plus lente que celle dun phénomène purement électrique.
A partir dun potentiel daction situé au point A dune fibre nerveuse, les régions adjacentes au point A sont au potentiel de repos (- 60 mV). La présence du potentiel daction (+ 50 mV) entraîne immédiatement la formation de courant dans les deux milieux intra et extra cellulaire. Lamplitude de ces courants est décrémentiel. Ces derniers vont traverser la membrane par différents points proches (B, C et D), lamplitude sera de plus en plus faibles au fur et à mesure que lon séloigne du point A, cette diminution est directement proportionnelle à la distance. Ceci soumet alors localement les protéines canal sensibles à la tension à un champ électrique plus ou moins important. Dans les régions où ces champs électriques dépassent le seuil douverture des protéines canal, lenchaînement des mécanismes de déclanchement dun potentiel daction en ce point est alors mis en jeu. Il apparaît alors au point C, quelques millisecondes plus tard, un nouveau potentiel daction et le cycle peut ainsi recommencer à partir de ce point.
Ce phénomène nest donc pas véritablement conduit le long de la fibre nerveuse mais régénérée de point en point. On qualifie ce phénomène de « propagation régénérative ».
La vitesse de propagation est plus rapide au sein des fibres de gros diamètres, ainsi quau sein des fibres myélinisées. Les cellules de Schwann forment une gaine de myéline isolante interrompu au niveau des nuds de Ranvier, ce sont les cellules elles même qui senroulent autour de laxone. Ces cellules jouent essentiellement un rôle dans la conduction saltatoire des potentiels daction. (Fig.7).

Les potentiels daction se reproduisent de place en place, lamplitude du phénomène, quel que soit lendroit où il apparaît est toujours le même. Cest ce que schématise le terme : tout ou rien. Il est nécessaire que chaque partie de la membrane possède des protéines canal spécifiques sensibles à la tension. Bien souvent ces protéines sont absentes des arborisations terminales. A ce niveau le potentiel daction va être transformé en signal chimique.
III. Potentiels post synaptiques excitateursUn même neurone reçoit des informations provenant de ces nombreux voisins par lintermédiaire de milliers de synapses dont certaines sont excitatrices et dautres inhibitrices. Le déclenchement du potentiel daction dans ce neurone dépend de sa capacité dintégrer ces informations multiples.
Dans une synapse excitatrice les récepteurs commandent un type de canaux chimio dépendant qui permet au Na+ dentrer dans la cellule et au K+ den sortir. Comme la force motrice du Na+ est supérieure à celle du K+, louverture de ces canaux produit un flux net de charges positives vers lintérieur de la cellule. Par conséquent la cellule se dépolarise et le potentiel de membrane se rapproche du seuil dexcitation, ce qui augmente la probabilité du déclenchement du potentiel daction par la cellule post synaptique. Le phénomène électrique provoqué par la liaison du neurotransmetteur spécifique au récepteur est appelé potentiel post synaptique excitateur ou PPSE. Le fait quun neurotransmetteur donné fasse apparaître un PPSE dépend du type de récepteur et de canaux chimio dépendant présent sur la membrane post synaptique. Le PPSE est un potentiel gradué dont lamplitude dépend du nombre de molécules de neurotransmetteur qui se lie au récepteur de la membrane post synaptique. La variation du potentiel électrique ne dure que quelques milli secondes, parce que les neurotransmetteurs sont dégradés par des enzymes. En outre, leffet électrique produit sur la cellule post synaptique décroît au fur et à mesure quon séloigne de la synapse. Pour que la cellule post synaptique soit activée, les courants ioniques locaux dus au PPSE doivent être assez fort pour que la dépolarisation de la membrane dans la région du cône dimplantation atteigne le seuil dexcitation. Le cône dimplantation est la zone de départ du potentiel de pointe, c'est-à-dire la région où les vannes des canaux tensio dépendant spécifiques aux Na+ souvrent pour générer un potentiel daction lorsquun stimulus dépolarise la membrane jusquau seuil dexcitation. Un PPSE du à une seule synapse, même proche du cône dimplantation, ne suffit habituellement pas à déclancher un potentiel daction. Cependant, plusieurs corpuscules nerveux terminaux agissant simultanément sur la même cellule post synaptique ou un nombre moins important de ces corpuscules libérant leur neurotransmetteur par rafale, peuvent avoir un effet cumulatif sur le potentiel de membrane à la hauteur du cône dimplantation. Cette addition des potentiels post synaptiques porte le nom de sommation.
Dans les deux cas la membrane na pas le temps de retrouver son potentiel de repos. Ainsi les courants ioniques peuvent dépolariser le cône dimplantation jusquau seuil dexcitation, activant le neurone (en déclanchant un potentiel daction). Les neurones effectuent donc des opérations complexes nécessitant la sommation de tous les PPSE pour produire une dépolarisation post synaptique significative : c'est l'intégration synaptique
Ces phénomènes élémentaires sont donc directement impliqués dans des systèmes de transmission de linformation au sein de lorganisme. Ils sont basés sur le potentiel de repos et les variations apportées permettent de créer un potentiel daction. Ce potentiel daction est reproduit identique à lui-même. Il est transformé sous forme chimique pour traverser les synapses, puis grâce aux potentiels post synaptique excitateurs il se recréé et se propage au sein de lorganisme.