{kind=link}
{kind=link}
{kind=link}
| Les hallucinogènes Psychedéliques |
Cette catégorie de drogue regroupe des excitants comme la mescaline (une phényléthanolamine), la psilocine (principe actif de la psilocibine générée par la dégradation de celle-ci) et le LSD (ces deux derniers étant des indolamines comme la 5HT).
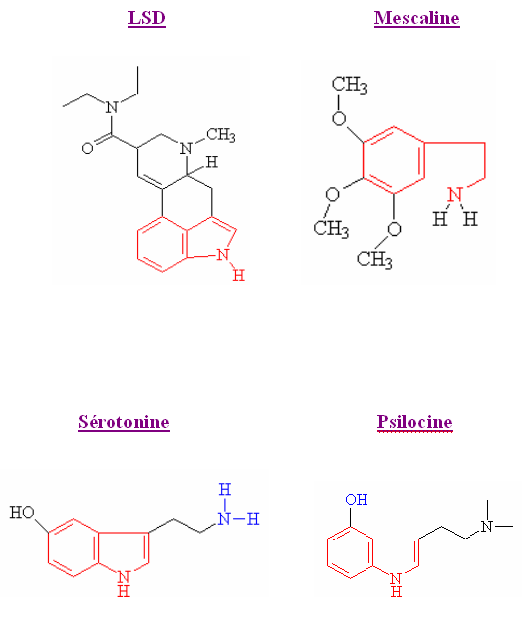
La psilocine est une forme hydrolysé où le groupe alcool a été changé de place.
En rouge : le noyau indol
En bleu : le groupe amine
Le LSD et la sérotonine ont deux anneaux de leur structure identiques et la chaîne latérale liée à ces anneaux de la sérotonine est similaire à une partie de la molécule de LSD. La structure de la mescaline est proche de celle de la noradrénaline ou de la dopamine et de la sérotonine.
Sous l'emprise de ces molécules, les utilisateurs ont l'impression d'être plus lucide et que les altérations sensorielles qu'ils ressentent reflètent une réalité supérieur à celle de l'état de conscience normale. Ces drogues révéleraient pour leurs prosélytes un niveau de perception surhumain. Finalement, c'est à cause de ces « mirages cognitifs » que l'on appelle ces drogues des psychédéliques, c'est à dire « qui manifeste l'esprit ».
Ces drogues entraînent donc d'importantes distorsions perceptuelles et cognitives recherchées par les toxicomanes. A ce titre et en toute rigueur, on ne devrait pas qualifier ces drogues d'hallucinogènes puisqu'elles ne créent pas de perceptions artificielles (comme le font les amphétamines et la cocaïne dans les psychoses et l'alcool dans le delirium tremens). Le LSD et ses analogues causent aussi des distorsions affectives ainsi que des troubles psychiatriques s'approchant de la schizophrénie. Aussi, devrait-on qualifier ces drogues de psychotomimétiques ou psychotogéniques : qui miment ou induisent des états psychotiques.
Des terreurs et des confusions mentales apparaissent sous l'effet de ces drogues. Mais l'une des plus graves séquelles de cette forme de toxicomanie est qu'elle déclenche des maladies mentales durables même lors de la première prise. En effet on connaît des centaines de cas de schizophrénie résultant d'un seul « bad trip ».
Dans le système nerveux central, les hallucinogènes produisent leurs manifestations psychédéliques en affectant le fonctionnement du locus coeruleus et les neurones glutamatergiques du néocortex par des mécanismes impliquant la neurotransmission sérotoninergique centrale.
I. Localisation des récepteurs sérotoninergiques.
Des études ont été faites avec plusieurs antagonistes de la 5HT marqué au 3H*, on a ainsi pu identifier les densités de récepteurs.
Pour les 2A, il y en a énormément dans les cortex préfrontal et temporal ; et quelques uns dans le cortex pariétal et les régions moteurs. A noter quil y en a très peu dans le thalamus.
Plus précisément les 5HT2A ont été identifiés en nombre dans le néocortex (couches III et V)
Pour les agonistes des amphétamines hallucinogènes les aires les plus denses en récepteurs sont les cortex frontal et claustrum, puis le noyau accubens et le bulbe olfactif.
Ce sont des agonistes qui se lient sur 5HT2A et 5HT2C. Les mRNa hybridation in situ sur le cortex humain ont montrés quil y a des 5HT2A sur les neurones pyramidaux et non pyramidaux.
Plus récemment, on a decouvert des 5HT2A dans les interneurones corticaux, en general les recepteurs sont situés sur une zone particulière du dendrite dans les régions extrasynaptique.
Ce sont les methodes dimmunomarquage et dautoradiographie qui ont permis de mettre en évidence ces récepteurs
Pour les 5HT1A qui sont des autorecepteurs, lhimunohistochimie a montré quil y aussi des 5HT1A dans lhippocampe, ceux ci auraient un rôle dans la modification du comportement.
II. Mode daction des hallucinogènes.Action sur les récepteurs 5HT1ALes structures moléculaires de la mescaline, de la psilocine et du LDS présentent des analogies avec celle de la sérotonine qui confèrent à ces drogues un potentiel sérotonino-mimétique. Ainsi le LSD et la psilocine sont des puissants inhibiteurs de l'activité électrique des neurones du raphé. L'inhibition est due à une action directe sur le soma des neurones sérotoninergiques du raphé où le LSD et la psilocine activent les autorécepteurs 5HT1A. Cependant, la seule activation des autorécepteurs 5HT1A est insuffisante pour le développement des hallucinations, car les anxiolytiques sérotoninergiques comme le buspirone agissent sur les autorécepteurs 5HT1A du raphé sans être hallucinogènes. D'autre part, la mescaline n'est qu'un faible agoniste des autorécepteurs 5HT1A mais elle est hallucinogène.
Action sur les récepteurs 5HT2
Par contre, tous ces psychédéliques ont en commun d'activer un autre type de récepteurs de la sérotonine : le récepteur 5HT2A (et plus particulièrement le sous type 5HT2A. A la différence des autorécepteurs 5HT1A présysnaptiques et inhibiteurs, les récepteurs 5HT2A sont postsynaptiques et excitateurs et ils sont présents dans de nombreuses structures cérébrales (dont les noyaux du tractus solitaire et accumbens) avec une forte concentration dans les neurones du néocortex.
Les hallucinogènes agissent via les récepteurs 5HT2A sur le néocortex et sur le locus coeruleus principalement. Ce dernier est le noyau regroupant l'essentiel des neurones noradrénergiques centraux (seulement 3000 neurones innervant 30 à 50% des neurones cérébraux). Il est constitué de 2 parties localisées dans le pont de Varole (mésencéphale) de part et d'autre du 4eme ventricule. Il reçoit des influx sensoriels, somatiques et viscéraux qu'il transmet à l'ensemble de l'encéphale grâce à ces axones très divergents. Les hallucinogènes inhibent les influx somatiques et viscéraux afférent au locus coeruleus, tout en facilitant les influx sensoriels. Cet effet double est réalisé par l'intermédiaire des récepteurs 5HT2A portés par des interneurones GABAergiques et glutamatergiques qui modulent le fonctionnement du locus coeruleus (voir schéma). Ainsi, la stimulation des interneurones GABAergiques augmente la libération de GABA dans le locus coeruleus ce qui inhibe, via des récepteurs GABA A, les influx somatiques et viscéraux. Inversement, l'activation des interneurones glutamatergiques renforce les influx sensoriels, par l'intermédiaire de récepteurs NMDA, et participe au développement des hallucinations. Il en résulte que le locus coeruleus envoie au néocortex des informations sensorielles altérées et amplifiées.
Au sein du néocortex, les psychédéliques perturbent la neurotransmission glutamatergique impliquant des interneurones qui établissent des synapses sur des neurones pyramidaux (neurones également glutamatergiques). Les interneurones possèdent des récepteurs 5HT2A sur leurs terminaisons qui modulent normalement, par des axones sérotoninergiques issus du raphé, la libération de glutamate. Les hallucinogènes activent les récepteurs 5HT2A, accroissant la libération de glutamate par les interneurones et stimulant ainsi les neurones pyramidaux dont l'activité devient alors anormale. La perturbation de la libération de glutamate par les neurones pyramidaux, dans le néocortex induits les hallucinations et les troubles cognitifs ainsi qu'une neurotoxicité glutamatergique nommée excitotoxicité. A cet effet direct sur le néocortex, s'ajoute la répercussion de l'activation du locus coeruleus par le LSD et ses analogues, car ses neurones libèrent de la noradrénaline sur des récepteurs alpha1 portés par les terminaisons des interneurones glutamatergiques. La stimulation des récepteurs noradrénergiques alpha1, accentue à l'instar des récepteurs 5HT2A la libération de glutamate sur les neurones pyramidaux.
III. Schéma récapitulatif.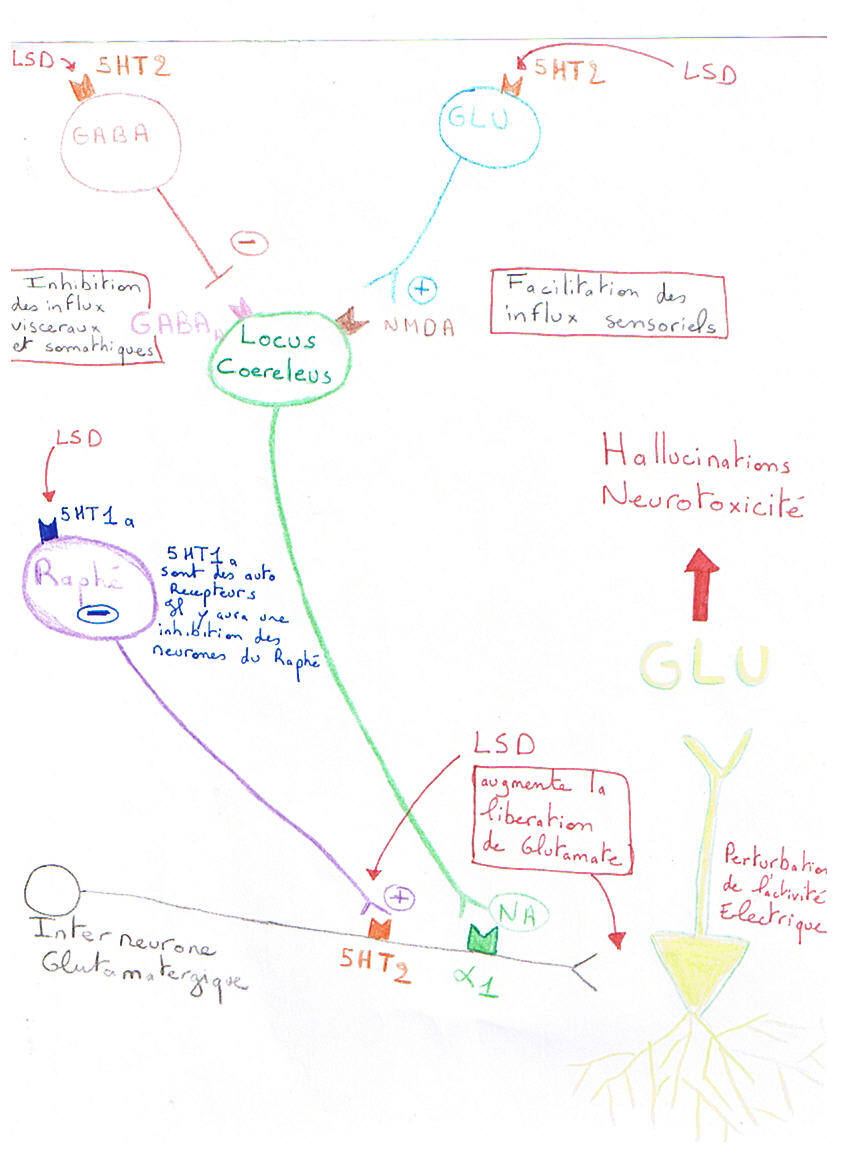
Les hallucinations sont dues à une perturbation de l'activité électrique des neurones pyramidaux du cortex frontal (néocortex). Cette perturbation résulte des effets des hallucinogènes sur le fonctionnement de plusieurs structures : le raphé, le locus coeruleus et le néocortex. Au niveau du raphé, les drogues inhibent l'activité neuronale en agissant sur les récepteurs 5HT1A ce qui modifie la libération de 5HT sur les interneurones glutamatergique du cortex. Dans le locus coeruleus, les hallucinogènes agissent sur les récepteurs 5HT2A et stimulent des interneurones gabaergiques, inhibant ainsi les influx somatiques et viscéraux. Par contre, les interneurones glutamatergiques du locus sont également activés car ils possèdent aussi des récepteurs 5HT2A; ces interneurones facilitent alors les influx sensoriels. Au total, le locus coeruleus envoie au cortex des informations sensorielles amplifiées et altérées ce qui se traduit par une élévation de la libération de noradrénaline (NA) sur les terminaisons des interneurones glutamatergiques du cortex. Dans le cortex, les hallucinogènes accroissent la libération de glutamate (neurotransmetteur excitateur) par les interneurones sur les neurones pyramidaux par une action directe sur les terminaisons dotées de récepteurs 5HT2A. A cette action directe, s'ajoute un effet indirect produit par la stimulation du locus coeruleus par les hallucinogènes. Cette stimulation accroît la libération de NA sur les terminaisons glutamatergiques, dotées de récepteurs adrénergiques alpha 1, et renforce la neurotransmission glutamatergique sur les neurones pyramidaux. Au bout du compte, la stimulation anormale des neurones pyramidaux altère leur fonctionnement ce qui produit les hallucinations et une neurotoxicité liée à une libération excessive de glutamante (excitotoxicité).